第1回 2019/4/9
- 講師:須谷 尚史
コース全体の説明
- 教科書:Molecular Biology of the Cell, 6th ed.
- 出席はとらない
- 成績はテストの結果のみ
- 出題範囲は教科書+授業で扱ったこと。
日程
| # | 月日 | 講師 | 内容 |
|---|---|---|---|
| 1 | 4/9 | 須谷 尚史 | 細胞周期研究の歴史概論 |
| 2 | 4/16 | 須谷 尚史 | DNA 複製の機構 |
| 3 | 4/23 | 須谷 尚史 | 染色体分配のメカニズム |
| 4 | 5/7 | 須谷 尚史 | 生殖細胞と減数分裂 |
| 5 | 5/14 | 須谷 尚史 | 細胞周期とチェックポイント |
| 6 | 5/21 | 須谷 尚史 | 染色体異常と発がん |
| 7 | 5/28 | 山中 総一郎 | 染色体構造とその制御 |
| 8 | 6/11 | 伊藤 耕一 | セントラルドグマ〜原核生物の転写装置・制御の基本 |
| 9 | 6/18 | 伊藤 耕一 | 真核生物の転写装置と高次な転写制御 |
| 10 | 6/25 | 山中 総一郎 | 染色体とエピジェネティクス |
| 11 | 7/2 | 伊藤 耕一 | 様々なmRNAの加工反応 |
| 12 | 7/9 | 伊藤 耕一 | 翻訳装置と遺伝暗号 |
| 13 | 7/16 | 伊藤 耕一 | 翻訳の分子機構とその制御 |
| 14 | 7/23 | 伊藤 耕一 | さまざまなRNA種と遺伝子発現制御 |
| 15 | 7/30 | 試験 |
細胞周期研究の歴史
| 内容 |
|---|
| 人間は Frog Development のような自律的な現象を作り出す事ができなかった。 |
| 物理などを考える前は、生気論(Vitalism)が中心。生き物は勝手に作られてるんだろう。(アリストテレス) |
| 自然発生説の否定:肉置いといても蛆虫わかない。親のハエがたからないと蛆虫湧かない |
| いや、でもそこに微生物おるやん!煮沸して中の微生物殺しても、ほっておくと微生物発生する。微生物が自然発生ちゃう??(これで200年間かかる) |
| 白鳥の首にすれば発生せん。ってことは空気中に微生物がいるだけやん!!(ルイ・パスツール) これがオートクレーブの元。 |
| その後、「尿素の合成(無機化合物から有機化合物を発生)」や「無細胞発酵(生物しかできないとされていた発酵を、死んだ生物でも実現した)」を通して、生命科学は化学の範疇なのかもしれないと考えられ始めた。 |
| 遺伝・複製(自分とよく似た子孫を残す)はどうやったら物理・化学的に説明する事ができるだろうか…? |
| メンデルの法則から「遺伝子」が、染色体の挙動がメンデルの法則に従うことから、「遺伝の染色体説」が広まる。 |
| シュレディンガー「生命現象は離散的であり、極めて量子的な性質を持つ。遺伝子は『非周期性の個体』であろう。」 |
| 1953年:DNAの分子構造の決定。二重螺旋構造をとっており、相補的なペアを作っているので、ある遺伝子から同じものをコピーする事が化学的に可能! |
| 遺伝子に注目が集まる。 |
| DNAの組み替え技術の発展により、生命科学研究が爆発的に進展した。 |
細胞周期 (Chapter.17)
細胞周期について
細胞は、次世代の細胞に遺伝情報を伝えるために、「染色体複製」と「染色体分配(細胞分裂)」という、倍にして分ける作業を繰り返している。なお、ほとんどの細胞は、通常ゲノムだけでなく細胞小器官や巨大分子も倍加するため、分裂のたびに小さくならないようになっている。

細胞周期の最も基本な機能は、「染色体の中のDNAを正確に倍加」してできた「コピーを正確に分離(染色体の分離・細胞分裂)」して遺伝的に同一な2つの娘細胞を作ることである。この2つの機能は非常に大事であり、細胞周期においてそれぞれS期(Synthesis)、M期(Mitosis)と呼ばれ、重要な時期を規定する。
ほとんどの細胞周期にはさらに間の時期がある。それぞれは内外の環境が上記の2つの期間に入るのに適切かを判断し、場合によっては進行を遅らせることで、前段階が未完了なまま周期の次の過程に進んだ場合に起こるトラブルを防ぐ。

細胞が細胞周期のどの段階にあるかの判断には、単純にDNA量を測る方法がある。これには、DNAと結合して蛍光を発する色素で細胞を染色し、蛍光量が細胞内のDNA量に直接比例するようにするという原理を利用したフローサイトメーター(FACS)が使われる。これによって、G1期、S期、G2期・M期の細胞の割合を求められ、各期間の長さが算定できる。

細胞周期制御系
疑問となるのは、どのように各期間を切り替えているのか、ということ。この役割は細胞周期制御系が果たしているのだが、ここでは、
- S期やM期のスイッチは何なのか?
- S期とM期を交互に繰り返すメカニズムはどのようなものか?
について解説していく。
サイクリン依存キナーゼ
細胞周期制御系の中心成分はサイクリン依存キナーゼ(cyclin-dependent kinase, Cdk)で、キナーゼ(リン酸化酵素)の活性が細胞周期の進行に伴って上下し、細胞周期の主要事象の開始や調節に関与するタンパク質のリン酸化状態を周期的に変化させる。
Cdk活性の周期的変動は、一連の複雑な酵素等のタンパク質の整然とした一連の複合体形成によって制御されている。Cdk調節因子の中で最も重要なのがサイクリン(cyclin)である。
Cdkは"dependent"という名前の通りサイクリンに依存し、サイクリンと強く結合していなければキナーゼ活性はない。なお、Cdkタンパク質の量は細胞周期で変化しないが、サイクリンタンパク質の周期的変化が、細胞周期の特定な段階でのサイクリン-Cdk複合体(cyclin-Cdk complex)の周期的な集合と活性化に繋がる。

サイクリンは、細胞周期に置いてCdkに結合し、機能する時期によって4種類に分類される。そのうち3種類(G1/S-cyclin, S-cyclin, M-cyclin)があらゆる真核細胞で必要とされる。

なお、酵母細胞では、1種類のCdkタンパクが細胞周期の各時期で結合するサイクリンを変え、細胞周期の様々な事象を促進するが、一方脊椎動物の細胞にはCdkが4種類あり、うち2つがG1-サイクリン、1つがG1/S-サイクリンとS-サイクリン、残りの1つがM-サイクリンに結合する。
これらは通例G1-Cdk, G1/S-Cdk, S-Cdk, M-Cdkと呼ばれる。
Cdk活性化キナーゼ
Cdkとサイクリンタンパクの三次元構造の研究により、サイクリンのないときにはCdkタンパクの活性部位が、タンパク質のループで一部塞がれていることが明らかになった。
サイクリンが結合するとそのループは取り払われ、Cdk酵素が部分的に活性化される。サイクリン-Cdk複合体の完全活性化には、別のキナーゼ、Cdk活性化キナーゼ(Cdk-activating kinase, CAK)によるCdk活性部位入り口付近のアミノ酸のリン酸化が必要である。

Cdk活性の抑制化
サイクリン量の増減の他にも、Cdk活性の調整にはいくつかの機構が存在する。
サイクリン-Cdk複合体の活性は、2個のアミノ酸のリン酸化によって阻害される。タンパクキナーゼWee1によるその部位のリン酸化がCdk活性を阻害し、ホスファターゼCdc25による脱リン酸化が活性を上昇させる。この両者による調節の仕組みが、有糸分裂の開始時点のM-Cdk活性の調節に特に重要である。

サイクリン-Cdk複合体はまた、Cdk阻害タンパク(Cdk inhibitor protein, CKI)の結合によっても調節される。

Cdk,Cyclin 発見の歴史
Cdkが見つかったのは、酵母遺伝学から。酵母は細胞サイズと形から、細胞周期の位相がわかるという観察上極めて重要な性質があった。

さらに、「温度感受性変異」という性質もあり、常温では色々な位相の細胞があるのに対し、高温にするとそれによって変異した遺伝子が本来働く時期に細胞周期が停止し、その時期の細胞のみを蓄積させることができた。

ここで、細胞周期が止まった細胞に色々な遺伝子を導入することで、高温での生育を救済するものを特定する研究がされた。そこで見つかったのが、MPF(M-phase promoting factor)であり、これが実はサイクリン-Cdk複合体であった;
MPF = Cdc2(Cdkキナーゼ) + Cdc13(サイクリン)
M-Cdk
ここでは、M期サイクリン複合体(M-Cdk)の活性を見ていく。なお、M-Cdkが有糸分裂のはじめに起こる多様で複雑な変化の全てを起動させているというのは驚きである。M-Cdkがリン酸化する標的タンパク質には、以下のようなものがある。
- コンデンシン(染色体凝縮)
- 微小管結合因子(紡錘体の形成)
- ラミン(核膜崩壊)
- ヒストン H1
G2→M
M-Cdkの活性化は、M-サイクリンの蓄積で始まる。胚ではM-サイクリンの合成は細胞周期を通じて一定で、間期にこのタンパク質が安定しているため、M-サイクリンが蓄積する。
この時M-サイクリンはCdk1と複合してM-Cdkの状態で蓄積しており、Cdk活性化キナーゼによって活性部位がリン酸化されているが、タンパクキナーゼ、Wee1による隣接部位2個の抑制性リン酸化のおかげで不活性化状態が保たれている。こうして細胞がG2期の終わりに達するころには、準備が整ったM-Cdkが豊富に貯蔵されながらも、その活性はキナーゼ活性部位をふさぐリン酸基により抑えられた状態になっている。
では、何がこの蓄積されたM-Cdkを活性化するのだろうか?ここで最も重要なのは、抑制性リン酸基を除去するタンパク質ホスファターゼ、Cdc25の活性化である。
この時、Cdc25もその標的であるM-Cdkによって部分的に活性化される。また、M-Cdkは抑制性キナーゼWee1を阻害する。M-Cdkが自身の活性化因子(Cdk25)を活性化し、阻害因子(Wee1)を阻害する能力を持つということは、有糸分裂におけるM-Cdkの活性化には正のフィードバックループが働くということを示唆している。これによって、G2期からM期への急激な遷移が引き起こされている。
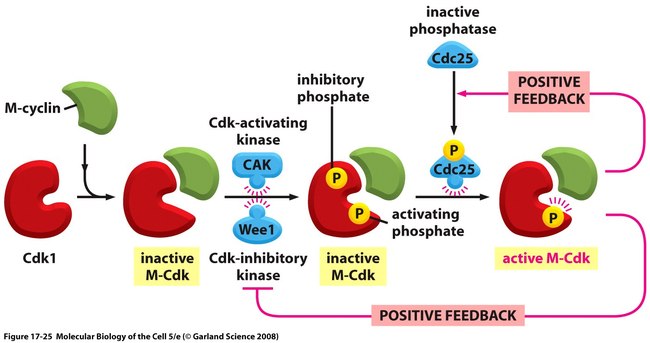
M→G1
M期からの脱出は、M-Cdk中のサイクリンが急激にタンパク質分解されることで引き起こされている。詳しくは説明しないが、後期促進複合体(APC/C)が標的タンパク質(サイクリン)にポリユビキチン鎖を付加し、それによって認識されるようになった標的がプロテアソーム内で分解される。
ここまでのまとめ
細胞周期の鍵となるCdk複合体は、
- リン酸化による活性制御
- タンパク質分解による不活性化
- 阻害タンパク質の結合
- (サイクリンの)転写制御
によって、その活性が制御されている。なお、今回はM期のみを見たが、M期以外の細胞周期においても、Cdk複合体が機能している。また、1つ前の段階が完遂されることが次の段階へ進む必須要件となっており、これによって細胞周期が正しい順序で進行するようになっている。

なお、主要な細胞周期調節タンパクを以下にまとめる。
| 一般名 | 機能と備考 |
|---|---|
| Cdkを修飾するタンパクキナーゼとタンパクフォスファターゼ | |
| Cdk活性化キナーゼ | Cdkの活性化部位をリン酸化 |
| Wee1キナーゼ | Cdkの抑制部位をリン酸化。有糸分裂前のCdk1の活性抑制に主に関与 |
| Cdc25フォスファターゼ | Cdkから抑制リン酸を除去。有糸分裂開始時にCdk1活性化の制御に主に関与 |
| ユビキチン連結酵素とその活性化因子 | |
| APC/C | セキュリンとSおよびM-サイクリンを含む主に有糸分裂の完了に関わる調節タンパクのユビキチン化を触媒。活性化タンパクCdc20またはCdh1の結合により調整される。 |
| Cdc20 | 全ての細胞のAPC/C活性化タンパク。中期-後期遷移におけるAPC/Cの最初の活性化を引き起こす。M-Cdc活性により活性化される。 |
哺乳細胞の細胞周期(特有)
- Rbタンパク質(図の赤色のタンパク質) (緑色はS期を引き起こす転写因子、G1期が終わったときにできる"hoge"(青色)が、Rbタンパク質を取り払い、それによってRbタンパク質が制御していた緑の転写因子がフリーになり、S期に進む。) なお、Rbはガン抑制因子で、それがない家系は網膜に腫瘍ができる事が知られている。(∵勝手に細胞周期が回るから。)
- ヒトパピローマウイルス(HPV)による細胞周期制御の破綻
- 増殖停止と再開

上から、「成長因子等の刺激」 → 「Mycの発現」 → 「G1-サイクリンの発現」といった流れ。理解が甘い。勉強せねば…
Glossary
| Words | Meanings |
|---|---|
| Cell, cycle | |
| G1, S, G2, M phases | |
| Cdk, Cyclin | |
| M-Cdk, S-Cdk, G1/S-Cdk | |
| Cdc25, Wee1, positive, feedback | |
| APC/C, protein, degradation | |
| G0 | |
| Rb, Myc |
Quizzes
| # | Question | Answer |
|---|---|---|
| 1 | What enzyme drives cell cycle progression into M phase? M期へと細胞周期を進める酵素は何か。 |
M-Cdk |
| 2 | What proteins compose this enzyme? その酵素の構成タンパク質は何か。 |
Cdk1 and Cyclin B (Vertebrates) ※種によって異なるが、ヒトの場合を覚えるのが一番安全かと。 |
| 3 | How does the enzyme promote M phase? その酵素はどのようにしてM期を開始するのか。 |
hosphorylates the substrates that function in mitotic cell cycle events. M期で何らかの機能を持つ基質(またはその上位にある酵素)をリン酸化する。 |
| 4 | How is the enzyme activated upon M phase onset? その酵素はM期の最初にどのようにして活性化されるか。 |
M-Cdk activity is kept low in G2 phase by inhibitory phosphorylation of Cdk1 by Wee1, Cdc25 dephosphorylates this inhibitory phosphorylation upon M phase onset, thereby activating M-Cdk. M-Cdk regulates Cdc25 positively and Wee1 negatively, which results in positive feedback and enables its rapid activation. M-Cdk は G1期にも存在するが、Wee1 によって抑制されている。 まず少量のM-Cdkができる。→ Cdc25活性化 → より多くのM-Cdk が脱リン酸化される(ポジティブフィードバック, 連鎖反応, 暴走的) |
| 5 | How is the enzyme inactivated upon M phase exit? その酵素はM期の最後にどのようにして不活性化されるか。 |
A M-Cdk subunit, Cyclin B, is degraded through targeted ubiquitylation catalyzed by APC/C CyclinBがAPC/Cのユビキチン化によって分解される。 |