第1回 2019/4/5
講義内容
- DNA・RNAの構造
- ゲノムの構造
- 遺伝暗号の縮重
- モデル生物におけるゲノムプロジェクト
- ヒトゲノムの遺伝子構成(次回)
イントロダクション
Gene sequencing cost analysis

| 年 | 内容 |
|---|---|
| 2000 | ドラフト配列の決定 |
| 2003 | ヒトゲノム計画終了 |
| 2007 | 次世代シークエンサーの誕生 |
ドラフト配列
10万塩基のクローンの配列決定を 10万塩基分 行った場合「カバー率1」という。
ここで、ゲノムの遺伝子地図を作成する際には、シークエンスした短い配列をつなぎ合わるため、確率論的に、正確な配列決定を行うためには10~20程度のカバー率が必要とされる。(この時の配列を完全配列と呼ぶ。)
ドラフト配列とは、全ゲノム配列に対して、カバー率 4~5で配列を決定したもののこと。
なお、次世代シークエンサー(リード長数十塩基)の場合は、ドラフト配列でもカバー率40~50が必要となっている。
Gene sequencing on 3 stages

| 手法 | 説明 |
|---|---|
| サンガー法 | DNA断片を持ってきて、蛍光のラベルを1つ1つ読んでいく。「末端でDNA伸長を止める」という技術を利用 |
| NGS | 数千万から数億のDNA断片に対して大量並列に一気に読む |
| 第三世代 | 反復配列などの、シークエンスするのが難しい配列を正しく読むことが目的。広い領域の配列を知る必要があるので、~kbp という量の塩基を読む。 |
Cost Per Genome
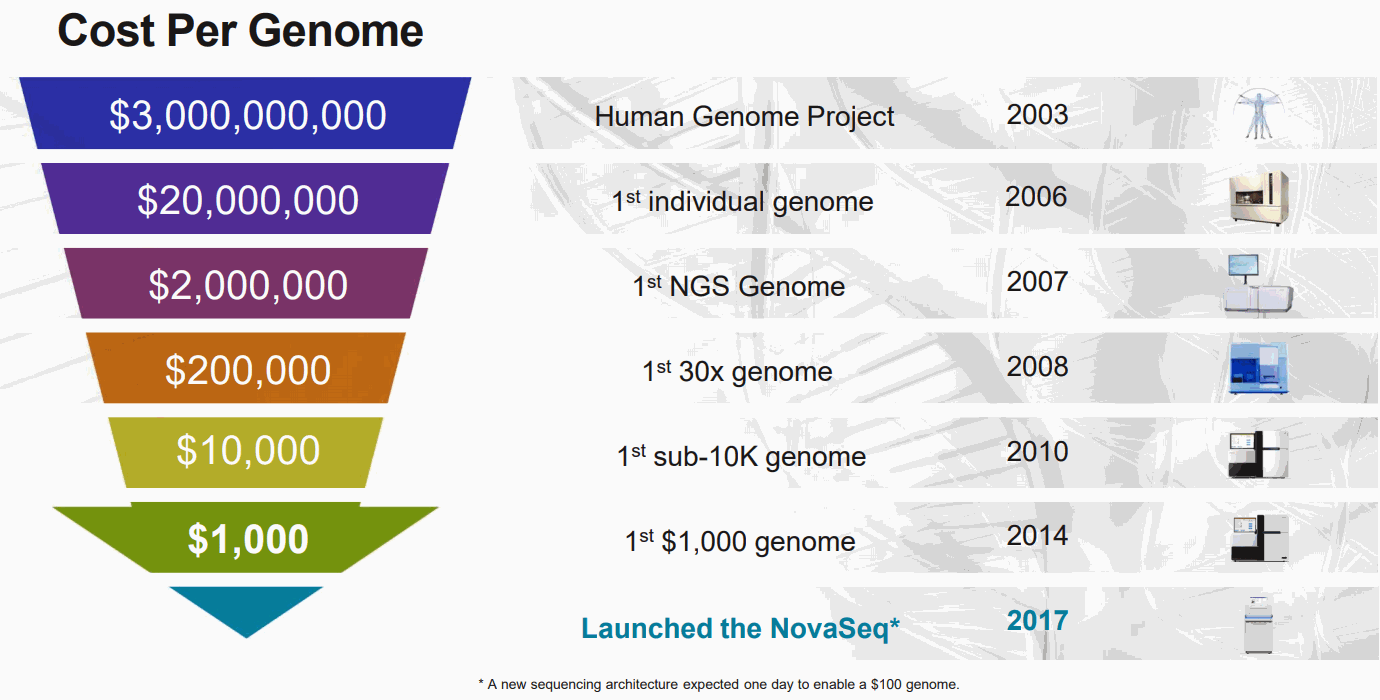
シークエンサーの進歩のおかげで、今や $1,000, 2days ぐらいで一人のゲノム解析ができるため、パーソナルゲノムの研究が現実的になっている。
Central Dogma
遺伝子の本体はDNAであり、それが転写されてRNAが作られ、3塩基のペアとアミノ酸が対応する。
この時、ただのアミノ酸配列では何もできず、適切な立体構造を作るというのが大事。
環境要因
双子のデータから「人の性格の50%は遺伝子に支配される」という報告もある。
↑ 性格の50%って何?🤔という感じ。どうやって定量評価しているのだろうか。
Genome(遺伝子) + Envirome(環境因子) = Phenome(表現系)
環境要因に対する応答
以下のように、実際に環境要因によって表現系に変化が出ているため、遺伝子の発現系のどこかで変化が起きているはずである。
| 生理的・生物学的要因 | 応答 |
|---|---|
| 光 | 光受容体タンパク質を介したシグナル伝達 |
| 放射線 | DNAの変異 |
| 電磁波 | DNAの変異 |
| 重力 | 骨量や筋肉現象を引き起こすエピゲノム変化(RNAの寄与?) |
| 湿度 | ? |
| 温度 | Gプロテインや温度センサーによる作用。RNAサイレンシングの制御 |
| ウイルス | RNAによるウイルスセンサーを介した作用 |
| 微生物 | RNAによる免疫受容体 |
| 寄生生物 | 腸内細菌や腸内におけるRNAの作用 |
| その他 | ヒトの場合、言語→思考・ストレス応答におけるRNA・エピゲノム変化? |
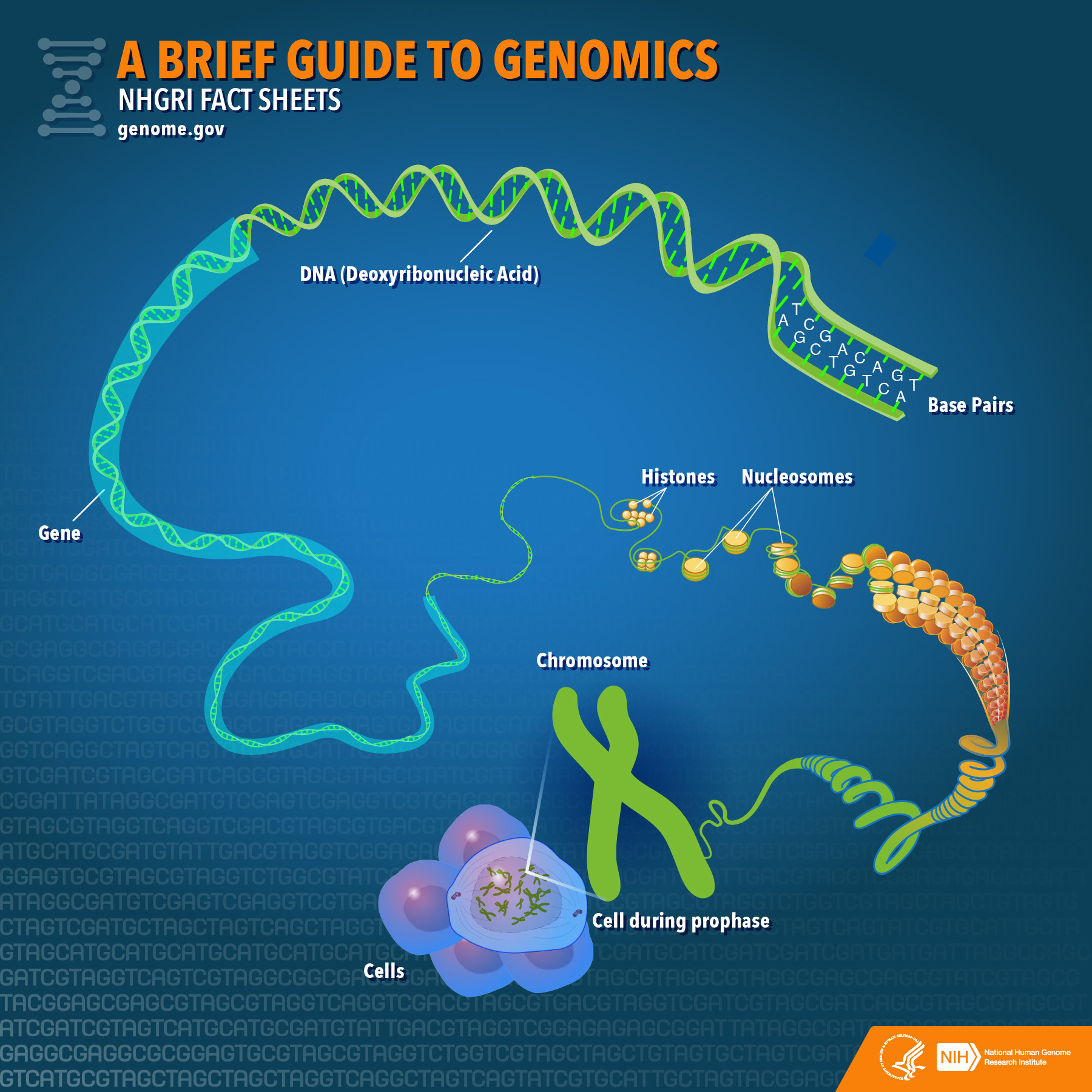
DNA・RNAの構造
- ヌクレオチド:[核酸塩基+糖+リン酸]
- ヌクレオシド:[核酸塩基+糖]

塩基・ヌクレオシド・ヌクレオチドの名称と略称

DNAとRNAの違い
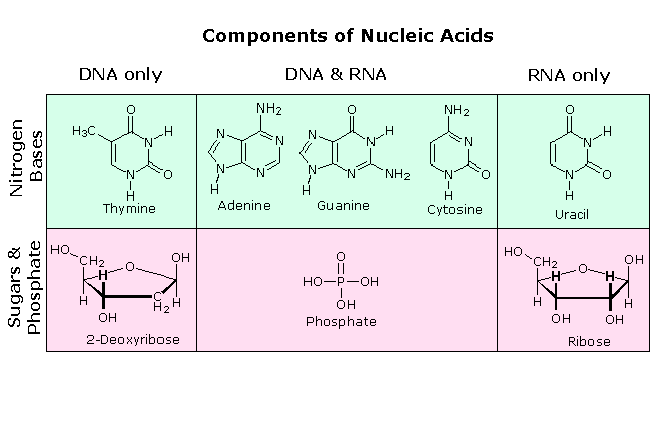
アデノシン・ADP・ATP

重合反応
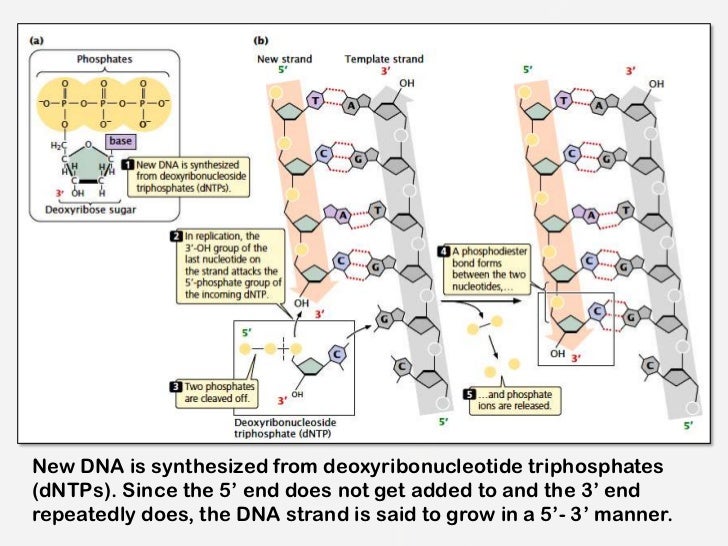
糖鎖がバッグボーンとなって結合が行われている。ここで、3番に記載があるように、monophosphateの状態で結合されている。
制限酵素を用いて特定のサイトで塩基配列を切ることはよくあるが、この時ホスホジエステル結合の切れ方には2種類ある(3'側には5'の炭素があり対称性がないから。)
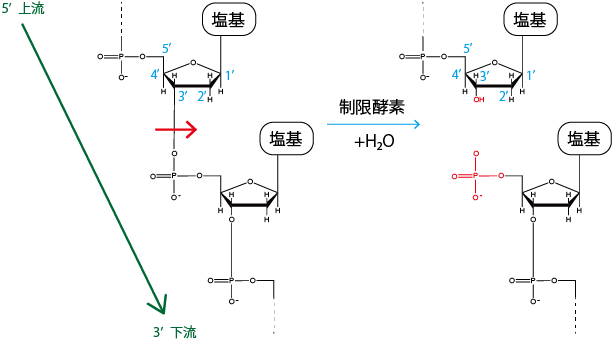
この二種類の切れ方を \(a\) 型 \(b\) 型と言い、それらがうまく繋がらないと結合できない。だから生物化学実験でEcoRⅠ処理したpUC4KとpUC119をライゲーションした時に方向が決まっていたのか!!
DNAが二重らせんを形成する根拠
- シャルガフ則: 各種DNA中のプリン、ピリミジンの存在比はほぼ1(∵ A/T=1, G/C=1)
- X線解析像
DNAの二重らせん
二重らせん構造の維持には以下の2つの特徴が重要である。
- 塩基対形成
- 塩基スタッキング DNA二重らせん中の塩基部分が重なり合っていること。電子を共有していると考えられており、これによって安定性を得ている。
Watoson-Crick塩基対
- プリンとピリミジンが対合する。
- TとAは2つの水素結合、CとGは3つの水素結合を形成する。
- 水素結合のエネルギー=1~10 kcal/molで、共有結合のエネルギーに比べると弱いが、数があるのでそれなりに強い。
例外
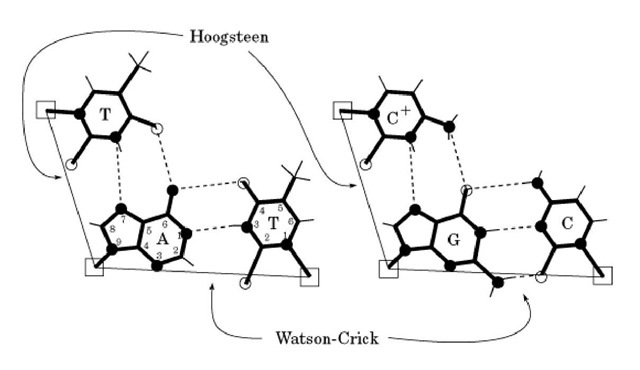
Hoogesteen塩基対
塩基対のパターンの一つであり、2つの核酸塩基が主溝に面した2本の水素結合によって結合する。天然で存在する割合はかなり低い(1%もない?)

三重鎖DNA
Watoson-Crick と Hoogesteen の両方を使うことで、DNAの三重鎖を形成することができる。
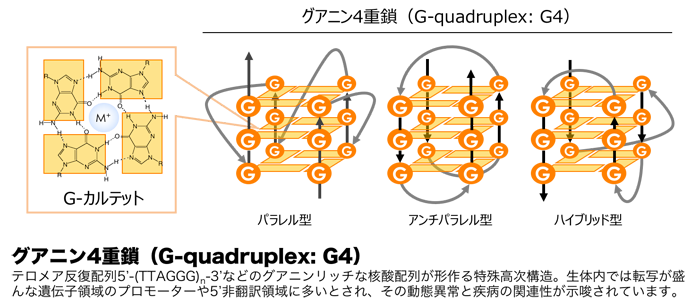
四重鎖DNA
Reverse Watson-Crick と Hoogesteen でG四重鎖ができる。この配列は、テロメアやプロモーターなどのG含有量の多い場所に現れる。
二重らせんの特徴
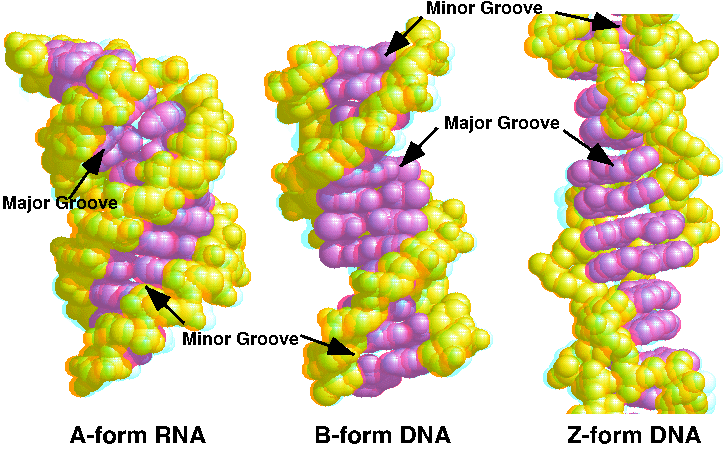
| A | B | Z | |
|---|---|---|---|
| 核酸 | RNA | DNA | DNA |
| らせんの構造 | 右巻き | 右巻き | 左巻き |
| 主溝のトポロジー(Major Groove) | 狭く深い | 広く深い | 平たい |
| 副溝のトポロジー(Minor Groove) | 広く浅い | 狭く浅い | 狭く深い |
ゲノムの構造
ヌクレオソーム
ヌクレオソームは真核生物の染色体構造の基本単位であり、ヒストンがないと秩序が崩壊する。

ヌクレオソームには4種類×2セット=8分子のヒストンからなるタンパク質のコア(芯)があり、円盤状のヒストン・コアの周りにDNAが左巻きにきつく1.7回転していることがわかっている。
DNAのたわみ
DNA二重らせんはヒストン八量体の周りに1.7回きつく巻きついているが、巻かれるためには内側が短く外側が長くなる必要があるので、GCの水素結合(3本)よりもATの水素結合(2本)の方が内側にきやすい(らせんの外側の方が強い力を受けるため?)。
このように、ヌクレオソーム・コアに対し親和性の高いDNA塩基配列があることが知られている。

遺伝暗号の縮合
RNAからタンパク質への情報の流れに伴う疑問
- 塩基対のような物理化学的な必然性はない。
- mRNAと合成中のタンパク質をつなぐアダプター分子が存在するのか?
- アミノ酸は一旦RNAと結合してからタンパク質に取り込まれる?
普遍暗号と非普遍暗号
基本的には(真核生物では)遺伝暗号表は普遍的であるが、種特異的な非普遍暗号を使う生物(細胞内小器官)が存在する。
遺伝暗号の縮重
1個のアミノ酸を複数のコドンが指定する。
20種類のアミノ酸(約20種類のアミノアシルtRNA合成酵素)に対し、tRNAは30~45種類(原核生物)、497種類、アンチコドン48種類(真核生物)。これに対してコドンは64(61)種類。
したがって、tRNAのアンチコドン48種類では、64(61)種類のコドンを識別することができない!
ゆえに、コドンの2番目までは正確な塩基対合をするが、3番目は誤っても大丈夫な"ゆらぎ(wobble)"を持つtRNAも存在する。